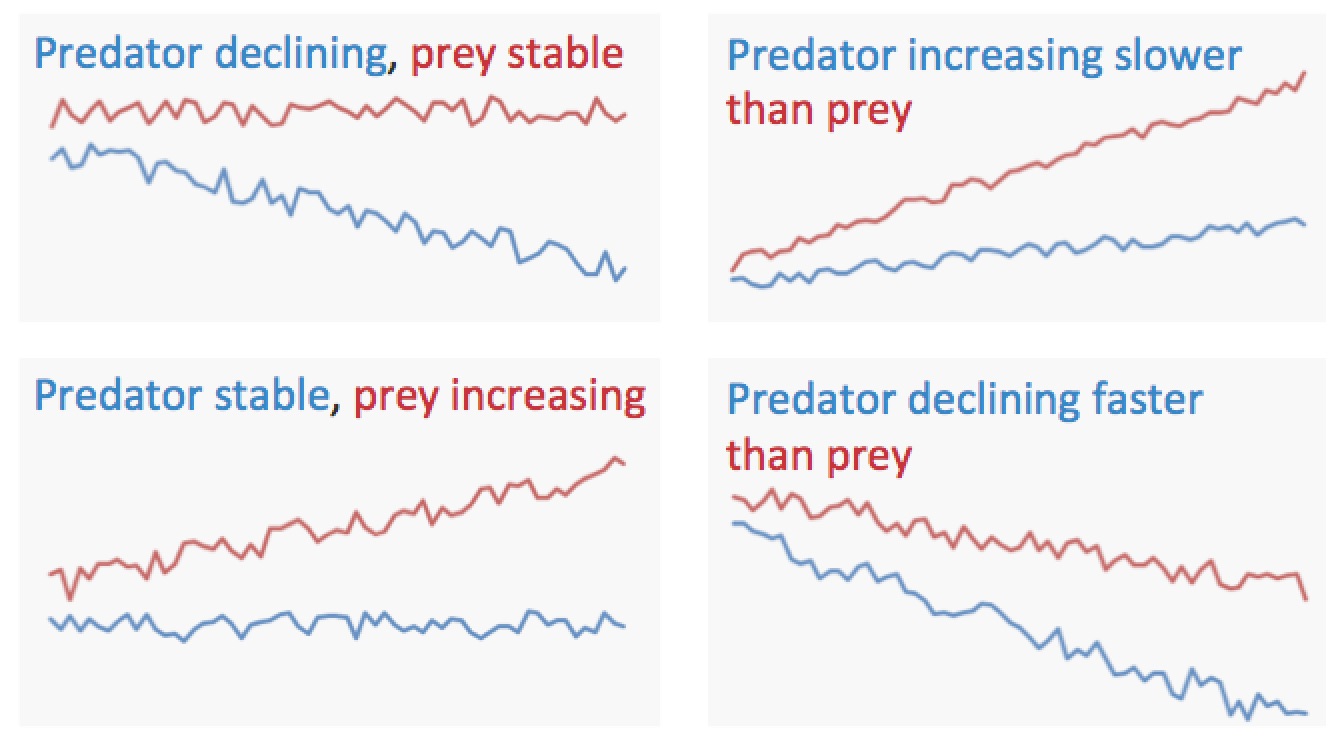
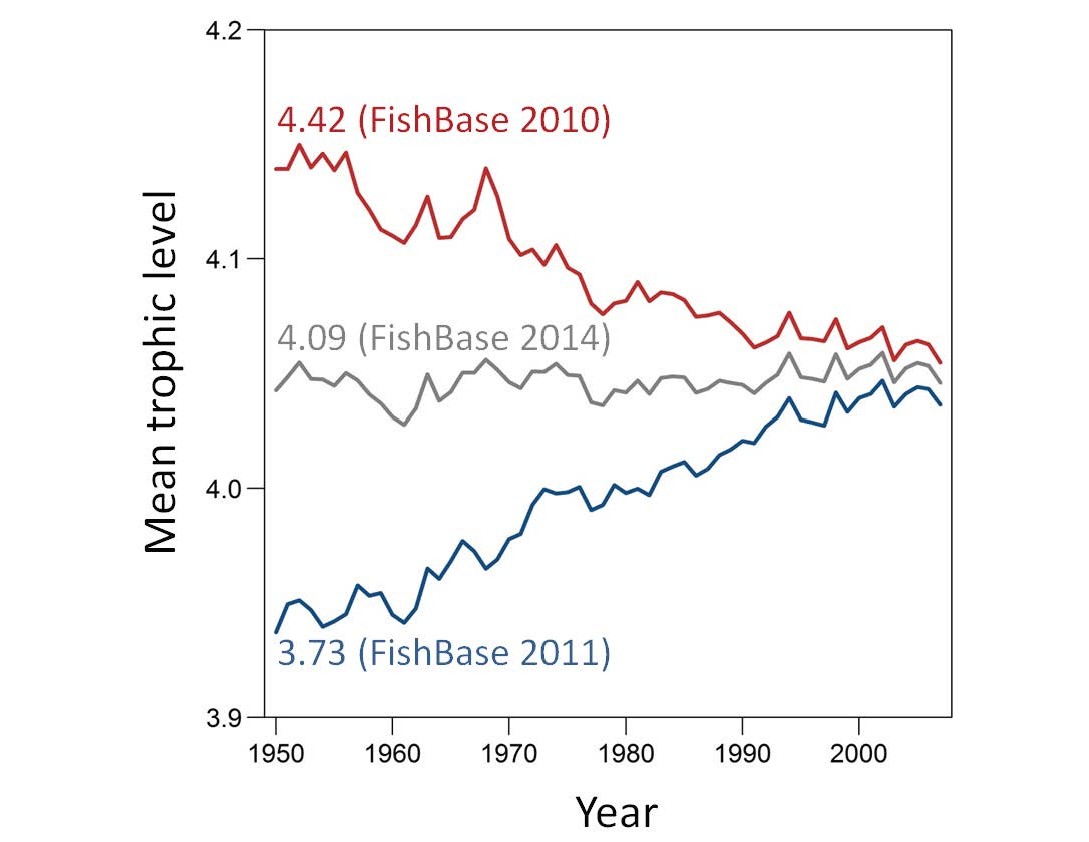
Trevor A. Branch School of Aquatic and Fishery Sciences, Box 355020, University of Washington, Seattle, WA, 98195. E-mail: tbranch@uw.edu Fisheries impact every part of marine food webs from the upper-trophic-level tunas and sharks to lower-trophic-level oysters and abalone. Although there are multiple ways in which fisheries develop, the dominant paradigm today is that we are “fishing down marine food webs”; first depleting top predators, then successively depleting species lower and lower in food webs (Christensen 1996; Pauly et al. 1998). The evidence for fishing down was declining mean trophic levels in global catch (Pauly et al. 1998), although declining mean trophic levels can result from several alternative scenarios (Figure 1), and mean trophic-level patterns in catches often do not match patterns in the ecosystem (Branch et al. 2010). Figure 1. Four scenarios of catches of predators (blue) and prey (red) that all result in declining mean trophic level. Declining mean trophic level can result from either fishing down marine food webs (a), (d), characterized by declining predator catches or from fishing through marine food webs (b), (c), when predator catches are stable or increasing, but catches of their prey expand over time. Regardless, new data show that mean trophic level of catch has steadily increased since the mid-1980s (Tacon and Metian 2009; Branch et al. 2010; Butchart et al. 2010; Tacon et al. 2010), unlike declines in all other global biodiversity indicators (Butchart et al. 2010). Additionally, if fishing down the food web is prevalent, top predators should be more likely to collapse, but data show that lower trophic-level species like abalone and oysters are more likely to collapse (Pinsky et al. 2011). Finally, there is no underlying economic reason for fishing down the food web: fisheries developed first not for predators but profits, targeting abundant, high-priced, and shallow-water species (Sethi et al. 2010). Thus, fishing down marine food webs is not occurring globally. There is evidence of fishing down in many ecosystems, although this evidence is confounded by studies that only report a decline in mean trophic level after adjusting data to exclude regions (Freire and Pauly 2010), remove large predators (Bhathal and Pauly 2008), exclude the most abundant species (Arancibia and Neira 2005), calculate declines only from years with declines (Bhathal and Pauly 2008), or remove all species below trophic level 3.25 or 3.5 (Pauly and Watson 2005; Pauly and Froese 2012). When tested at a global scale, removing all species below 3.5 initially resulted in a decline in mean trophic level (Branch et al. 2010; Pauly and Froese 2012); this turned into an increase (Branch 2012) when the Atlantic Cod Gadus morhua trophic level was updated in FishBase and has subsequently turned into a stable trend following the most recent Atlantic Cod update (Figure 2). Figure 2. Trends in mean trophic level in global catch for species above trophic level 3.5, modified from Branch et al. (2010) to take into account changes in the estimated trophic level for Atlantic Cod. The trophic-level estimate for Atlantic Cod in FishBase (Froese and Pauly 2014) changed from 4.42 in 2010 to 3.73 in 2011, reverted to 4.42 in 2012–2013, and changed again to 4.09 in 2014. Recently, the argument for fishing down has shifted to fisheries expansion as the primary reason why declining mean trophic level in catch has not yet been detected in all areas of the world (Pauly and Palomares 2005; Bhathal and Pauly 2008; Kleisner et al. 2014; Grüss 2015). The basic premise is that fisheries first fished down low-trophic-level nearshore species and then expanded to offshore predators like tunas, and this is argued to mask the evidence for fishing down. Fisheries expansion has certainly occurred (Morato et al. 2006; Swartz et al. 2010), but shifting from low to high trophic levels over time is evidence for fishing up marine food webs, not fishing down marine food webs. This is confirmed by trends in mean trophic level by bins of latitude and distance from the coast (figure 3 in Kleisner et al. 2014), which over 1985 to 2004 show increases for 74% of all bins, with more increases than decreases at every distance from the coast. I assert that instead of focusing on one hypothesis about how fishing affects marine food webs, we should be following the method of multiple working hypotheses (Chamberlin 1965). Chamberlin (1965:755) warns that “The search for facts, the observation of phenomena and their interpretation, are all dominated by affection for the favored theory until it appears to its author or its advocate to have been overwhelmingly established. The theory then rapidly rises to the ruling position.” To avoid the “blinding influence” of such affection, Chamberlin urges that multiple working hypotheses be kept in mind when weighing evidence. Instead of focusing on one hypothesis about how fishing affects marine food webs, we should be following the method of multiple working hypotheses. In this instance, there are at least five working hypotheses available to explain how fishing affects food webs, which are not mutually exclusive and may vary in space and time: 1. Fishing down marine food webs (Christensen 1996; Pauly et al. 1998). 2. Fishing up marine food webs: fishing first targets lower trophic levels and then shifts to predators (Essington et al. 2006; Erlandson et al. 2009; Litzow and Urban 2009). 3. Fishing through marine food webs: catches on top predators stay high or increase, but fishing also expands on lower trophic levels (Essington et al. 2006). 4. Balanced exploitation: fishing expands over time on all species, regardless of trophic level (Branch et al. 2010; Zhou et al. 2010; Garcia et al. 2012). 5. Fishing for profits: fisheries developed based on profit rather than trophic levels, first targeting abundant, highly priced, and easily accessible species, before expanding to other less profitable species (Sethi et al. 2010). To reliably assess how fisheries affect marine food webs, we must first lay out plausible hypotheses, then examine the evidence for each, and finally decide which hypotheses can be discarded and which ones remain. Following this approach, Essington et al. (2006) found that fishing down occurs in 9 ecosystems, fishing up in 18 ecosystems, and fishing through in 21 ecosystems. As Chamberlin (1965:759) put it, we need “… to balance all available evidence fairly, and to accept that interpretation to which the weight of evidence inclines, not that which simply fits our working hypothesis or our dominant theory.” Moreover, by probing the circumstances that favor particular hypotheses, we are likely to identify which management actions result in different impacts of fishing on marine food webs. REFERENCES Arancibia, H., and S. Neira. 2005. Long-term changes in the mean trophic level of central Chile fishery landings. Scientia Marina 69:295–300. Bhathal, B., and D. Pauly. 2008. “Fishing down marine food webs” and spatial expansion of coastal fisheries in India, 1950–2000. Fisheries Research 91:26–34. Branch, T. A. 2012. FAO’s state of fisheries and aquaculture: correcting some misrepresentations by Pauly and Froese. Marine Policy 36:1191–1192. Branch, T. A., and coauthors. 2010. The trophic fingerprint of marine fisheries. Nature 468:431–435. Butchart, S. H. M., M. Walpole, B. Collen, A. van Strien, J. P. W. Scharlemann, R. E. A. Almond, J. E. M. Baillie, B. Bomhard, C. Brown, J. Bruno, K. E. Carpenter, G. M. Carr, J. Chason, A. M. Chenery, J. Csirke, N. C. Davidson, F. Dentener, M. Foster, A. Galli, J. N. Galloway, P. Genovesi, R. D. Gregory, M. Hockings, V. Kapos, J.-F. Lamarque, F. Leverington, J. Loh, M. A. McGeoch, L. McRae, A. Minasyan, M. H. Morcillo, T. E. E. Oldfield, D. Pauly, S. Quader, C. Revenga, J. R. Sauer, B. Skolnik, D. Spear, D. Stanwell-Smith, S. N. Stuart, A. Symes, M. Tierney, T. D. Tyrrell, J.-C. Vié, and R. Watson. 2010. Global biodiversity: indicators of recent declines. Science 328:1164–1168. Chamberlin, T. C. 1965. The method of multiple working hypotheses. Science 148:754–759 (original publication date 1890). Christensen, V. 1996. Managing fisheries involving predator and prey species. Reviews in Fish Biology and Fisheries 6:417–442. Erlandson, J. M., T. C. Rick, and T. J. Braje. 2009. Fishing up the food web?: 12,000 years of maritime subsistence and adaptive adjustments on California’s Channel Islands. Pacific Science 63:711– 724. Essington, T. E., A. H. Beaudreau, and J. Wiedenmann. 2006. Fishing through marine food webs. Proceedings of the National Academy of Sciences U.S.A. 103(9):3171–3175. Freire, K. M. F., and D. Pauly. 2010. Fishing down Brazilian food webs, with emphasis on the east Brazil large marine ecosystem. Fisheries Research 105(1):57–62. Froese, R., and D. Pauly, editors. 2014. FishBase. World Wide Web electronic publication. Available: www.fishbase.org. (July 2014). Garcia, S. M., and J. Kolding, J. Rice, M.-J. Rochet, S. Zhou, T. Arimoto, J. E. Beyer, L. Borges, A. Bundy, D. Dunn, E. A. Fulton, M. Hall, M. Heino, R. Law, M. Makino, A. D. Rijnsdorp, F. Simard, and A. D. M. Smith. 2012. Reconsidering the consequences of selective fisheries. Science 335:1045–1047. Grüss, A. 2015. Q&A: the present and the future of world and U.S. fisheries—interview with Daniel Pauly. Fisheries 40(1):37–41. Kleisner, K., H. Mansour, and D. Pauly. 2014. Region-based MTI: resolving geographic expansion in the Marine Trophic Index. Marine Ecology Progress Series 512:185–199. Litzow, M. A., and D. Urban. 2009. Fishing through (and up) Alaskan food webs. Canadian Journal of Fisheries and Aquatic Sciences 66:201–211. Morato, T., R. Watson, T. J. Pitcher, and D. Pauly. 2006. Fishing down the deep. Fish and Fisheries 7(1):24–34. Pauly, D., V. Christensen, J. Dalsgaard, R. Froese, and F. Torres, Jr. 1998. Fishing down marine food webs. Science 279:860–863. Pauly, D., and R. Froese. 2012. Comments on FAO’s State of Fisheries and Aquaculture, or “SOFIA 2010.” Marine Policy 36:746–752. Pauly, D., and M.-L. Palomares. 2005. Fishing down marine food web: it is far more pervasive than we thought. Bulletin of Marine Science 76(2):197–211. Pauly, D., and R. Watson. 2005. Background and interpretation of the “Marine Trophic Index” as a measure of biodiversity. Philosophical Transactions of the Royal Society of London B 360:415–423. Pinsky, M. L., O. P. Jensen, D. Ricard, and S. R. Palumbi. 2011. Unexpected patterns of fisheries collapse in the world’s oceans. Proceedings of the National Academy of Sciences U.S.A. 108:8317– 8322. Sethi, S. A., T. A. Branch, and R. Watson. 2010. Fishery development patterns are driven by profit but not trophic level. Proceedings of the National Academy of Sciences U.S.A. 107:12163–12167. Swartz, W., E. Sala, S. Tracey, R. Watson, and D. Pauly. 2010. The spatial expansion and ecological footprint of fisheries (1950 to present). PLOS ONE 5(12):e15143. Tacon, A. G. J., and M. Metian. 2009. Fishing for aquaculture: nonfood use of small pelagic forage fish—a global perspective. Reviews in Fisheries Science 17(3):305–317. Tacon, A. G. J., M. Metian, G. M. Turchini, and S. S. de Silva. 2010. Responsible aquaculture and trophic level implications to global fish supply. Reviews in Fisheries Science 18:94–105. Zhou, S., A. D. M. Smith, A. E. Punt, A. J. Richardson, M. Gibbs, E. A. Fulton, S. Pascoe, C. Bulman, P. Bayliss, and K. Sainsbury. 2010. Ecosystem-based fisheries management requires a change to the selective fishing philosophy. Proceedings of the National Academy of Sciences U.S.A. 107:9485–9489.
Recent News
This website uses cookies to improve your experience. AcceptRead More
Privacy & Cookies Policy
Privacy Overview
This website uses cookies to improve your experience while you navigate through the website. Out of these, the cookies that are categorized as necessary are stored on your browser as they are essential for the working of basic functionalities of the website. We also use third-party cookies that help us analyze and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these cookies. But opting out of some of these cookies may affect your browsing experience.
Necessary cookies are absolutely essential for the website to function properly. This category only includes cookies that ensures basic functionalities and security features of the website. These cookies do not store any personal information.
Any cookies that may not be particularly necessary for the website to function and is used specifically to collect user personal data via analytics, ads, other embedded contents are termed as non-necessary cookies. It is mandatory to procure user consent prior to running these cookies on your website.